|
La conductance à la
vapeur d'eau d'un oeuf actuel ou fossile (GH2O ,
mg H2O torr / jour ) est établie en utilisant la
longueur effective moyenne des canaux aérifères
( L, cm ) et la surface porale totale (estimation Ap, cm2 ).Elle est établie selon
l'équation suivante :
GH2O = 23,42 x Ap
x L-1 ( Seymour , 1990 )
La conductance à la
vapeur d'eau d'un oeuf avien est estimée selon
l'équation : GH20 = 0,432 x M 0.78 ( où M =
masse de l'oeuf ( g ) ( Ar et alii ,
1974 ).La conductance d'un grand nombre d'oeufs
aviens actuels a été établie par Tullett (1978).
Sur la base d'un
important matériel ( 66 oeufs entiers ou
partiellement entiers collectés par nos soins
depuis 1975 ) nous avons révisé les résultats
obtenus par Williams , Seymour et
Kerourio ( 1984 , tabl.II ) concernant
Megaloolithus aureliensis , Cairanoolithus
dughii , M. siruguei , M. mamillare .Nous
avons , par ailleurs , établi la conductance de
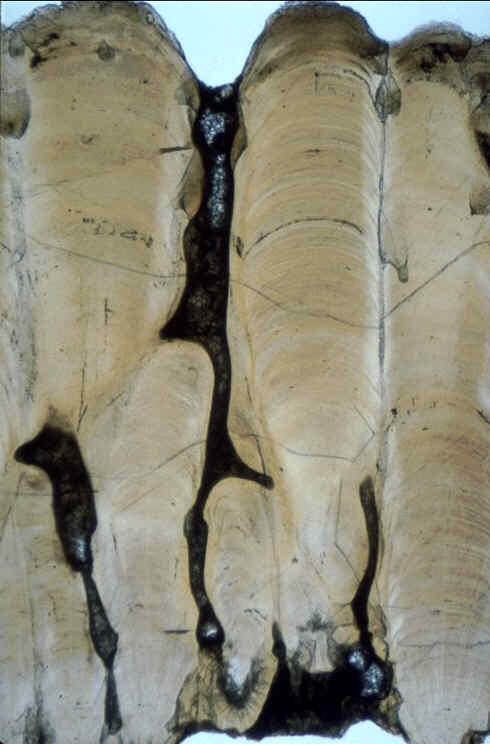
la coquille à la vapeur d'eau pour
Prismatoolithus matellensis et trois
ootypes inédits.
La conductance des
types d'oeufs étudiés est de 8 à 87 fois
supérieure à celles calculées pour des oeufs
aviens de mêmes dimensions .Ces nouvelles
observations confirment partiellement les
données obtnues par Seymour ( 1980 ), Seymour &
Ackerman ( 1980 ) et Williams , Seymour et
Kerourio ( 1984 ).
La très haute
conductance des différents types d'oeufs
fossiles présents dans le bassin
d'Aix-en-Provence résulte essentiellement de
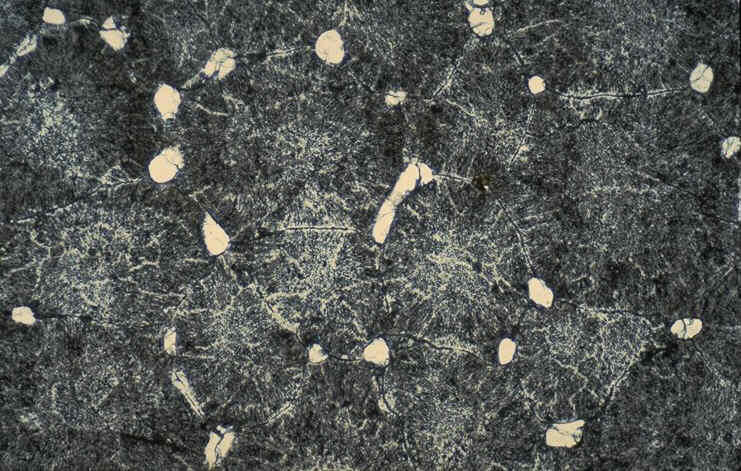
l'importance des surfaces porales ( de 9 à 79
cm 2 suivant les ootypes ).L'extension
inhabituelle des surfaces porales résulte du
nombre très important de pores (de 100 000 à
près de 600 000 selon les ootypes ), soit de 3 à
13 fois le nombre de pores prédit pour des oeufs
aviens de masse identique selon l'équation :
1499 x masse de l'oeuf ( g ) puissance 0,420 (
Tullett & Board , 1978 ).
Les oeufs à
coquille rigide déposés par les reptiles actuels
nidifiant dans des milieux très humides ont des
conductances variant de 2 à 5,5 fois celles
d'oeufs aviens de dimensions équivalentes .Les
valeurs calculées pour des oeufs aviens à
coquille souple sont encore plus élevées (Tracy
et alii, 1978 ; Packard et alii
, 1979 ; Packard et alii , 1982 ; Lutz
et alii , 1980 ; Ackerman et alii
, 1985 ; Seymour & Ackerman , 1980 ; Ar &
Rahn , 1985 ).
Ces résultats
confirment l'hypothèse établie sur la base des
données paléontologiques , mais souvent
contreversées , selon lesquelles les oeufs
fossiles du Maastrichtien du bassin
d'Aix-en-Provence étaient incubés dans des
conditions de haute humidité et dans un
environnement très probablement souterrain (
Seymour , 1980 ; Seymour & Ackerman , 1980 ;
Williams ,Seymour et Kerourio, 1984 ;
Kerourio , 1987 ). Inversement l'hypothèse d'une
nidification aérienne apparaît
physiologiquement improbable , et même
fantaisiste. En milieu aérien la perméabilité
des coquilles aurait été trop élevée et , si
les exigences de l'embryon en oxygène auraient
été satisfaites , il aurait aussi perdu trop
d'eau et il se serait rapidement déshydraté .Il
est évident que les conditions d'incubation de
la plupart des oeufs aviens actuels auraient été
léthales aux embryons dinosauriens (Kerourio ,
1987 ).La thèse d'une nidification aérienne (Dughi
et Sirugue , 1958 et 1976 ; Breton et alii
, 1986 a et b et 1994 ) nous semble reposer sur
une interprétation inadéquate des données
paléontologiques et sur une méconnaissance
complète de la physiologie de l'oeuf reptilien.
|
Types d'œufs
|
Megaloolithus
aureliensis |
Cairanoolithus dughii |
Megaloolithus siruguei |
Megaloolithus
mamillare |
ootype 1 |
ootype 2 |
ootype 3 |
|
Surface porale estimée
( cm2 ) |
20,4 |
14 |
32,8 |
16,1 |
79,1 |
9,81 |
53,2 |
|
Longueur moyenne d'un
pore ( en cm ) |
0,09 |
0,19 |
0,18 |
0,09 |
0,1 |
0,07 |
0,14 |
|
GH2O estimé (mg / torr
/ jour) |
5137,2 |
1707,7 |
4244 |
4236,6 |
18903,2 |
3429,1 |
8899,6 |
|
GH2O prédit avien |
173,5 |
223 |
236,8 |
256,5 |
216,1 |
80,9 |
147,4 |
|
Rapport GH2O prédit à
GH2O avien |
29,6 |
7,6 |
17,9 |
16,5 |
87,4 |
42,3 |
60,3 |
Porosité de la
coquille et conductance à la vapeur d'eau pour
quelques ootypes du Maastrichtien continental du
bassin d'Aix-en-Provence ( Bouches-du-Rhône ,
France ). |